The Host Immune Response in Wound Infection
Wound healing involves a series of highly coordinated and overlapping phases, which includes an inflammatory phase, a proliferative phase, and a remodeling phase.1 In an acute wound, these phases occur in a timely manner;2-4 however, many chronic wounds become stalled in the inflammatory phase of healing whereby excess inflammatory cells together with elevated levels of pro-inflammatory cytokines and proteases persist within the wound tissue.5,6 This creates a hostile wound environment causing tissue damage. Furthermore, inflammatory cells deplete the wound of oxygen required for effective tissue repair.7 Infection amplifies the immune response, increasing wound chronicity.8
The host responds to infection by upregulating numerous pro-inflammatory cytokines.8 Neutrophils are the primary immune cell type to respond to these signals, arriving at the wound site and releasing oxidative and proteolytic enzymes.9 Neutrophils engulf and digest the microorganisms and are responsible for removing foreign material, digesting necrotic tissue, and producing cytokines to support the proliferative phase of healing.10 Monocytes, which differentiate into macrophages, along with dendritic cells and mast cells, also respond to infection to support the innate immune response.11 Once bioburden is controlled, macrophages digest apoptotic neutrophils to prevent excess tissue damage caused by neutrophils, and signal to resolve the inflammatory phase of healing.12 This innate immune response is highly effective in healthy individuals; however, comorbidities can disrupt this system preventing the effective control of wound bioburden.13
The Impact of Wound Infection
Wound infection increases patient morbidity and mortality and represents a major financial burden to healthcare providers. A recent UK study by Guest and colleagues estimated that “59% of chronic wounds healed if there was no evidence of infection compared to 45% if there was a definite or suspected infection”.14 Furthermore, 80-100% of hospital admissions attributable to venous or mixed aetiology leg ulcers, diabetic foot ulcers or open wounds were linked to suspected infection.14 The impact that wound infection has on patient morbidity and mortality and the financial burden it puts on healthcare providers clearly merits greater research into the biological response to infection, and infection prevention and treatment strategies.
An introduction to biofilms, definitions, and prevalence in chronic wounds.
The wound bed provides a moist, warm, and nutritious environment, whereby microorganisms can acquire nutrients from necrotic tissue or plasma and lysed red blood cells leaked from capillaries.15,16 This environment can support the formation of biofilm within the wound.
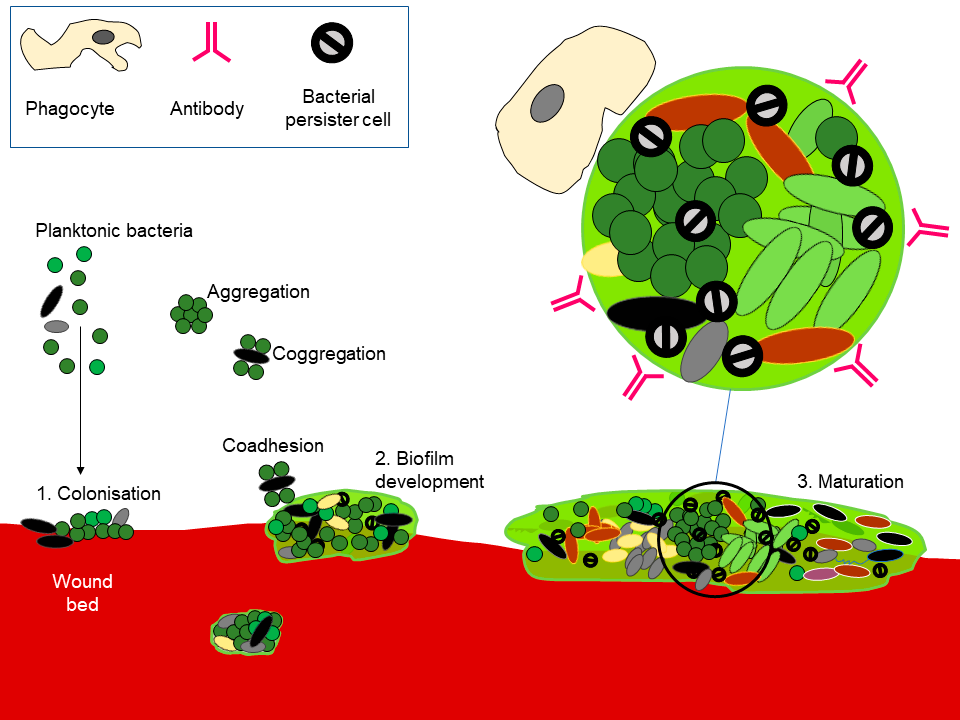
The wound bed can be colonized by microorganisms, as aggregates or in dispersed, planktonic form (1). Given time, and in the absence of effective interventions and immune response, biofilms can develop (2). Exogenous microorganisms can potentially integrate into a formed biofilm (3) or can be removed by physical disruption. Image courtesy of Andrew McBain, B.Sc, PhD.
Microbial populations can be defined based on the arrangement of cells as planktonic (where cells are dispersed, often within a liquid) or as biofilms. Whilst some definitions of biofilms extend into any situation where microorganisms interact with surfaces, recalcitrance, or antimicrobial tolerance, a key feature of biofilms, is strongly associated with the presence of an extracellular matrix composed principally of polymeric material that, in wounds, may originate both from the microorganisms and the host.17 Biofilm formation requires microorganisms, water, nutrients, often a surface for colonization, and time. In practice, biofilms can form in many moist environments including those with limited nutrient availability. The nutrient-rich environment of wounds can be highly permissive of biofilm development. Microorganisms are generally more amenable to control with antimicrobials before they have formed or integrated into a biofilm and it has been shown that susceptibility to antimicrobials decreases rapidly on attachment to a surface and this tolerance is likely to increase as the matrix develops.18,19 Figure 1 illustrates some aspects of biofilm formation that apply to recalcitrant wound infections. As tolerance to antibiotics is a key biofilm characteristic,20 it is necessary to differentiate between infections where biofilms are likely to be aetiologically important and those where this is unlikely to be the case.
Biofilms have been frequently reported in chronic wound samples and have been implicated in the chronicity of chronic infections. Evidence for the presence of biofilms in chronic wounds comes from observational studies. Malone conducted a meta-analysis of published literature and reported the overall prevalence of biofilms in chronic wounds as 78.2% (CI 61.6–89, p<0.002) but was no lower than 60% in any individual study, with many of the studies reporting 100% biofilm prevalence.24 By contrast, a lower biofilm prevalence has been reported in acute wounds. For example, a highly cited paper by James and colleagues assessed chronic wound specimens and acute wound specimens reporting that “of the 50 chronic wound specimens evaluated by microscopy, 30 were characterized as containing biofilm (60%), whereas only one of the 16 acute wound specimens was characterized as containing biofilm (6%).”25
The tolerance of biofilms to the host immune response and antimicrobial therapies is believed to represent a significant problem in wound healing.17 Researchers have noted two major facets of delayed wound healing:26,27
i) The “broken host” theory, which proposes that once a breach in the skin barrier occurs, the composed host environment facilitates bacterial colonization that has a neutral effect on the wound healing process;
ii) That the microbiota represents a major barrier to healing in chronic wounds, suggesting that chronic wounds are chronic infections frequently involving biofilms.28,29 A key requirement for biofilm formation is time, and this is more abundant in non-healing wounds.30
Microorganisms can colonize and/or infect the wound bed resulting in delayed healing. However, both host and microbial factors are likely to be important, with a complex interplay between both factors likely.13
Kalan and colleagues conducted a longitudinal, prospective study of patients with neuropathic diabetic foot ulcers reporting that “strain-level variation of Staphylococcus aureus and genetic signatures of biofilm formation were associated with poor outcomes”.31 Similarly, Corynebacterium striatum isolated from wounds, and applied to full-thickness excisional wounds in a diabetic mouse model of impaired wound healing was associated with an early delayed healing phenotype.31
In summary, most studies indicate that biofilms are present in the majority of chronic wounds. These may be principally bacterial but can comprise multiple species of microorganism variously arranged in aggregates. Evidence is accumulating to implicate wound biofilms in poor prognosis, but more research is needed to better understand the importance of the taxonomic composition of the wound microbiota to prognosis.
The Host Response to Biofilms
Despite infected wounds exhibiting a heightened state of inflammation, the host immune response does not effectively combat biofilm infection. Observational studies have shown that neutrophils surround the biofilm but are unable to penetrate it and kill the microorganisms within.32 Ultimately, this protection results in an excess neutrophil accumulation within the wound. In addition, in vitro studies have shown that macrophage response to biofilms exhibit limited phagocytosis capabilities and are deficient in the expression of alternatively activated M2 markers.33
Although many studies have shown that biofilms heighten the immune response, other studies have reported that biofilms are no more virulent than planktonic infection.34 Sweere and colleagues reported no difference in proinflammatory cytokine expression between wounds infected with planktonic or pre-formed biofilms suggesting the early inflammatory response is similar between planktonic and biofilm wound infections.34
Co-morbidities increase the risk of wound infection. In diabetic patients, a dysregulated immune response predisposes patients to infection. Biofilm infected wounds in diabetic mice exhibit significantly less proinflammatory cytokines and TLR2 and TLR4 expression compared to biofilm infected wounds in wild-type mice.6 Although biofilm infected wounds of diabetic and wild-type mice exhibit a similar level of bioburden and infiltration of neutrophil after three days, in diabetic wounds, neutrophil oxidative burst activity is reduced. This results in a significant increase in bioburden after ten days and a delay in healing.6
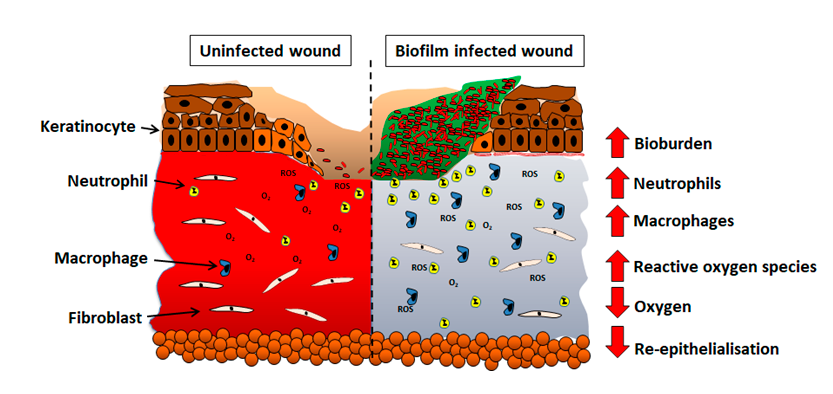
The host response of persistent inflammation to biofilm infection has subsequent impacts on the later phases of healing. During normal tissue repair, the recruitment of inflammatory cells is tightly regulated to prevent excess tissue damage; however, in biofilm infected wounds, excess inflammation results in tissue damage. Roy and colleagues reported that in the remodeling phase of healing, biofilm infections influence collagen turnover within the granulation tissue.35 An increase in collagen degradation correlates with an increase in matrix metalloproteases which regulate the turnover of collagen during healing. Biofilm infections have also been shown to delay wound re-epithelialization to a greater extent than planktonic infections36 and this may occur through the inhibition of keratinocyte binding to fibronectin receptors in the matrix altering keratinocyte migration,37 or as a secondary effect due to increased inflammation. These animal studies highlight the ineffectiveness of the host immune response at combating biofilm infection and the effects it has on the healing of biofilm infected wounds. Figure 2 illustrates changes in the wound environment in biofilm infected wounds. Thus, in a clinical setting, therapeutic strategies are required to aid the host immune response in biofilm resolution.
Biofilm Tolerance and Strategies to Combat it.
Tolerance towards antimicrobial compounds in biofilms has been an area of active research for many decades.38-41 This tolerance can be readily demonstrated in the laboratory with biofilms often having and exceeding 1000 times less susceptibility than the same microorganisms grown in dispersed, planktonic mode.42,43 Such tolerance will frequently render a biofilm untreatable with systemic antibiotics.44 It is commonly believed that cellular aggregation, high cell density and the extracellular matrix are key factors in the protection of biofilms against both antimicrobials and immune factors, as illustrated in Figure 3.18,19,22,23
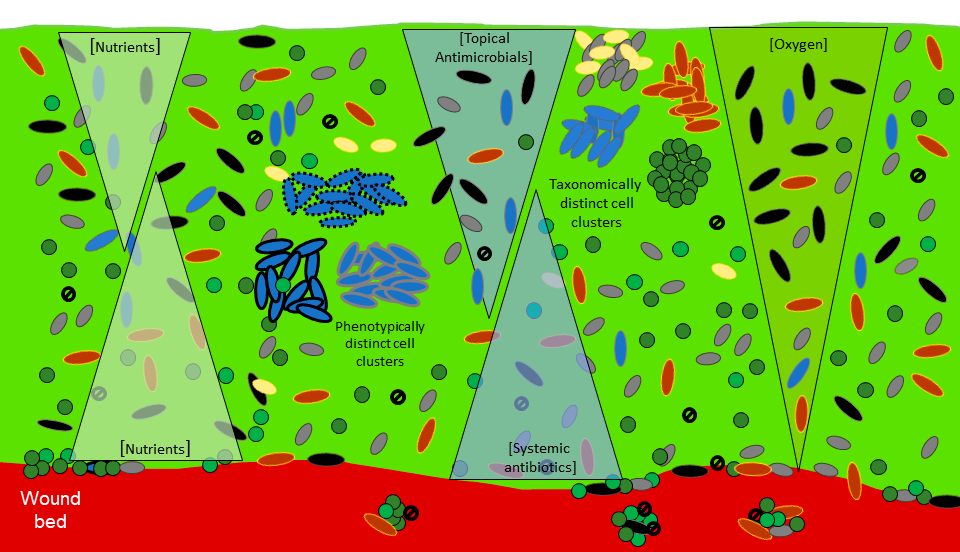
The extracellular matrix is a key biofilm feature in many environments including wounds.17 Whilst evidence suggests that the extracellular matrix is important in conferring protection to biofilms against antimicrobials this is not primarily due to simple protection from penetration.47-50 The process is distinct from pharmacological antibiotic resistance; is reversible, in that disruption of a biofilm where it can be achieved, can result in susceptible bacteria; and often resembles a Pyrrhic victory in that tolerance may be driven by the regrowth of survivors post-treatment.18 The description of biofilm tolerance to antimicrobials as a multilayered defence,51 and multicellular resistance captures key elements of the mechanisms involved.52 The following points reflect important considerations for the deployment of antimicrobials during wound management: